
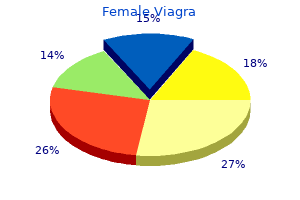
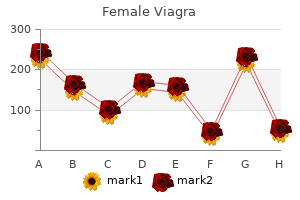
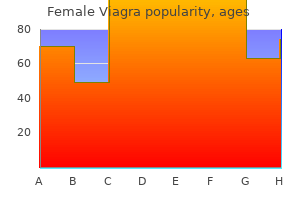
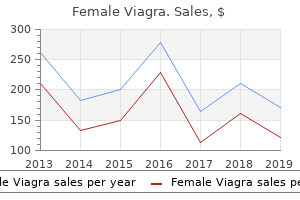
Female Viagra
"Generic 50mg female viagra fast delivery, womens health medical group fort worth".
By: D. Ugolf, M.A., M.D.
Assistant Professor, Weill Cornell Medical College
When the plasma glucose is high pregnancy leg cramps best purchase for female viagra, insulin secretion is normally increased and hepatic glucogenesis is decreased women's health clinic campbelltown female viagra 100mg free shipping. Glucagon also contributes to the hyperglycemia pregnancy zone protein effective female viagra 50mg, and glucose output by the liver is facilitated by catecholamines pregnancy jokes cartoons order female viagra with mastercard, cortisol, and growth hormone when the stress of illness is severe. Effects of Hyperglycemia Hyperglycemia by itself can cause symptoms resulting from the hyperosmolality of the blood. In addition, there is glycosuria because the renal capacity for glucose reabsorption is exceeded. Excretion of the osmotically active glucose molecules entails the loss of large amounts of water (osmotic diuresis; see Chapter 38). The resultant dehydration activates the mechanisms regulating water intake, leading to polydipsia. Increasing the oral caloric intake to cover this loss simply raises the plasma glucose further and increases the glycosuria, so mobilization of endogenous protein and fat stores and weight loss are not prevented. The role of chronic hyperglycemia in production of the long-term complications of diabetes is discussed below. Effects of Intracellular Glucose Deficiency the plethora of glucose outside the cells in diabetes contrasts with the intracellular deficit. Glucose catabolism is normally a major source of energy for cellular processes, and in diabetes, energy requirements can be met only by drawing on protein and fat reserves. Mechanisms are activated that greatly increase the catabolism of protein and fat, and one of the consequences of increased fat catabolism is ketosis. Deficient glucose utilization in the cells of the hypothalamic ventromedial nuclei is probably the cause of the hyperphagia in diabetes. When the activity of the satiety area of this nucleus is decreased in response to decreased glucose utilization in its cells, the lateral appetite area operates unopposed, and food intake is increased (see Chapter 14). Glycogen depletion is a common consequence of intracellular glucose deficit, and the glycogen content of liver and skeletal muscle in diabetic animals is usually reduced. This shift is indicated in Figure 19-10, which also shows the other principal abnormalities of intermediary metabolism in the liver. Some idea of the rate of gluconeogenesis in fasting diabetic animals is obtained by measuring the ratio of glucose (dextrose) to nitrogen in the urine (D/N ratio). In fasting animals, liver glycogen is depleted and glycerol is converted to glucose at a very limited rate, so that the only important source of plasma glucose is protein (see Chapter 17). It can be calculated that the amount of carbon in the protein represented by 1 g of urinary nitrogen is sufficient to form 8. Consequently, the D/N ratio of approximately 3 seen in diabetes indicates the conversion to glucose of about 33% of the carbon of the protein metabolized. Glucagon stimulates gluconeogenesis, and hyperglucagonemia is generally present in diabetes. Adrenal glucocorticoids also contribute to increased gluconeogenesis when they are elevated in severely ill diabetics. There is an increased supply of amino acids for gluconeogenesis because, in the absence of insulin, less protein synthesis occurs in muscle and hence blood amino acid levels rise. In addition, the activity of the enzymes that catalyze the conversion of pyruvate and other two-carbon metabolic fragments to glucose is increased. These include phosphoenolpyruvate carboxykinase, which facilitates the conversion of oxaloacetate to phosphoenolpyruvate (see Chapter 17). They also include fructose 1,6-diphosphate, which catalyzes the conversion of fructose diphosphate to fructose 6-phosphate, and glucose 6-phosphatase, which controls the entry of glucose into the circulation from the liver. Increased acetyl-CoA increases pyruvate carboxylase activity, and insulin deficiency increases the supply of acetyl-CoA because lipogenesis is decreased. Pyruvate carboxylase catalyzes the conversion of pyruvate to oxaloacetate (see Figure 17-9). Protein depletion from any cause is associated with poor "resistance" to infections. Fat Metabolism in Diabetes the principal abnormalities of fat metabolism in diabetes are acceleration of lipid catabolism, with increased formation of ketone bodies, and decreased synthesis of fatty acids and triglycerides. The manifestations of the disordered lipid metabolism are so prominent that diabetes has been called "more a disease of lipid than of carbohydrate metabolism.
One such toxin is batrachotoxin women's health best body meal plan reviews purchase generic female viagra on line, produced by a species of frog; some tribes of South American Indians use this poison on their arrow tips pregnancy 30 weeks cheap 50 mg female viagra overnight delivery. A number of plants produce similar toxins women's health clinic eau claire wi purchase female viagra 50mg overnight delivery, including aconitine women's health clinic parramatta buy cheap female viagra 100 mg, from buttercups; veratridine, from lilies; and a number of insecticidal toxins produced by plants such as chrysanthemums and rhododendrons. Peptide toxins affecting K+ channels include dendrotoxin, from wasps; apamin, from bees; and charybdotoxin, yet another toxin produced by scorpions. All of these toxins block K+ channels as their primary action; no toxin is known to affect the activation or inactivation of these channels, although such agents may simply be awaiting discovery. These "naked" ions are then able to move through four K+ binding sites within the selectivity filter to eventually reach the extracellular space (recall that the normal concentration gradient drives K+ out of cells). On average, two K+ ions reside within the selectivity filter at any moment, with electrostatic repulsion between the two ions helping to speed their transit through the selectivity filter, thereby permitting rapid ion flux through the channel. Crystallographic studies have also determined the structure of the voltage sensor in another type of bacterial K+ channel. Such studies indicate that the sensor is at the interface between proteins and lipid on the cytoplasmic surface of the channel, leading to the suggestion that the sensor is a paddle-like structure that moves through the membrane to gate the opening of the channel pore (Figure 4. Crystallographic work has also revealed the molecular basis of the rapid transitions between the closed and the open state of the channel during channel gating. By comparing data from K+ channels crystallized in what is believed to be closed and open conformations (Figure 4. Producing a "kink" in one of these helices increases the opening from the central water-filled pore to the intracellular space, thereby permitting ion fluxes. These paddles reside within the lipid bilayer of the plasma membrane and may respond to changes in membrane potential by moving through the membrane. The gating charges that sense membrane potential are indicated by red "plus" signs. Opening of the pore of the channel involves kinking of a transmembrane domain at the point indicated in red, which then dilates the pore. The best-characterized of these diseases are those that affect skeletal muscle cells. In these disorders, alterations in ion channel proteins produce either myotonia (muscle stiffness due to excessive electrical excitability) or paralysis (due to insufficient muscle excitability). Other disorders arise from ion channel defects in heart, kidney, and the inner ear. Channelopathies associated with ion channels localized in brain are much more difficult to study. Nonetheless, voltage-gated Ca2+ channels have recently been implicated in a range of neurological diseases. These include episodic ataxia, spinocerebellar degeneration, night blindness, and migraine headaches. Red regions indicate the sites of these mutations; the red circles indicate mutations. Usually, attacks are precipitated by emotional stress, exercise, or alcohol and last for a few hours. Abnormal retinal function may arise from decreased Ca2+ currents and neurotransmitter release from photoreceptors (see Chapter 11). This defect has been mapped to two mutations: one on chromosome 2 that encodes an subunit for a voltage-gated Na+ channel, and the other on chromosome 19 that encodes a Na+ channel subunit. This disease is characterized by frequent brief seizures commencing within the first week of life and disappearing spontaneously within a few months. A reduction in K+ current flow through the mutated channels probably accounts for the hyperexcitability associated with this defect. Mu- tant channels inhibit the function of other, non-mutant K+ channels and may produce clinical symptoms by impairing action potential repolarization. Mutations in the K+ channels of cardiac muscle are responsible for the irregular heartbeat of patients with long Q-T syndrome. Numerous genetic disorders affect the voltage-gated channels of skeletal muscle and are responsible for a host of muscle diseases that either cause muscle weakness (paralysis) or muscle contraction (myotonia).
Auditory Responses of Neurons in the Medulla Oblongata the response of individual second-order neurons in the cochlear nuclei to sound stimuli are like those of the individual auditory nerve fibers menstruation while nursing order female viagra with mastercard. The frequency at which sounds of the lowest intensity evoke a response varies from unit to unit; with increased sound intensities menstruation in the middle ages cheap female viagra 50mg online, the band of frequencies to which a response occurs becomes wider women's health clinic andrews afb buy female viagra 100mg fast delivery. The major difference between the responses of the first- and second-order neurons is the presence of a sharper "cutoff" on the low-frequency side in the medullary neurons breast cancer knee socks order female viagra 100mg amex. This greater specificity of the second-order neurons is probably due to some sort of inhibitory process in the brain stem, but how it is achieved is not known. Primary Auditory Cortex the pathways from the cochlea to the auditory cortex are described in the first section of this chapter. Impulses ascend from the dorsal and ventral cochlear nuclei through complex paths that are both crossed and uncrossed. In animals, there is an organized pattern of tonal localization in the primary auditory cortex, as if the cochlea had been unrolled upon it. In humans, low tones are represented anterolaterally and high tones posteromedially in the auditory cortex. However, it is pitch and not frequency per se that is coded in the auditory cortex, because when a complex sound with a missing fundamental is presented (see above), the part of the cortex that is stimulated is the part corresponding to the perceived pitch. Thus, processing of pure frequencies into pitch must occur at a subcortical level. The auditory pathways in the cortex resemble the visual pathways in that there is increasingly complex processing of auditory information along them, and there is even emerging evidence that like the visual pathways, they are organized in two general paths. In the case of the auditory system, there seems to be a dorsal-parietal pathway concerned with sound localization (the "where" pathway). The parietal areas are concerned, for example, with sound movement, and, interestingly, there are neurons in the frontal cortex of monkeys that respond only to near sounds, the sounds which obviously have the greatest survival importance to the animal. On the other hand, the human temporal lobes contain a ventral "what" pathway, including areas where the neurons respond selectively to voices. These voice-selective areas may be analogous to the face-selective areas associated with the visual system (see Chapter 8). Like the somasthetic and visual cortical systems, the auditory system is modified by experience and other factors. The increase in the size of auditory receiving areas produced by auditory stimuli paired with high-frequency stimulation of the basal forebrain pathways in rats was mentioned in Chapter 7. Examples of auditory plasticity in humans include the observation that in individuals who become deaf before language skills are fully developed, viewing sign language activates auditory association areas outside the primary auditory cortex. Conversely, individuals who become blind early in life are demonstrably better at localizing sound than individuals with normal eyesight. In other studies, babies were found to rapidly develop enhanced neuronal responses to sounds unique to their native language after 6 months of age, whereas responses to sounds that were not unique gradually disappear. In these individuals, there is an increase in the size of the auditory areas activated by musical tones. In addition, violinists have altered somatosensory representation of the areas to which the fingers they use in playing their instruments project. Musicians also have larger cerebellums than nonmusicians, presumably because of learned precise finger movements. A portion of the posterior superior temporal gyrus known as the planum temporale (Figure 9-16) is regularly larger in the left than in the right cerebral hemisphere, particularly in right-handed individuals. A curious observation which is presently unexplained is that the planum temporale is even larger than normal on the left side, ie, the asymmetry is greater, in musicians and others who have perfect pitch. Sound Localization Determination of the direction from which a sound emanates in the horizontal plane depends upon detecting the difference in time between the arrival of the stimulus in the two ears and the consequent difference in phase of the sound waves on the two sides; it also depends upon the fact that the sound is louder on the side closest to the source. The detectable time difference, which can be as little as 20 us, is said to be the most important factor at frequencies below 3000 Hz and the loudness difference the most important at frequencies above 3000 Hz. Neurons in the auditory cortex that receive input from both ears respond maximally or minimally when the time of arrival of a stimulus at one ear is delayed by a fixed period relative to the time of arrival at the other ear. Sounds coming from directly in front of the individual differ in quality from those coming from behind, because each pinna (the visible portion of the exterior ear) is turned slightly forward. In addition, reflections of the sound waves from the pinnal surface change as sounds move up or down, and the change in the sound waves is the primary factor in locating sounds in the vertical plane. This device presents the subject with pure tones of various frequencies through earphones.
The concentration of Ca2+ in the outer segment appears to play a key role in the light-induced modulation of photoreceptor sensitivity young women's health birth control pills discount female viagra 50 mg fast delivery. This decrease triggers a number of changes in the phototransduction cascade pregnancy journal online buy female viagra from india, all of which tend to reduce the sensitivity of the receptor to light womens health jackson ms purchase 100 mg female viagra mastercard. The regulatory effects of Ca2+ on the phototransduction cascade are only one part of the mechanism that adapts retinal sensitivity to background levels of illumination; another important contribution comes from neural interactions between horizontal cells and photoreceptor terminals (see below) womens health center xenia ohio buy genuine female viagra on line. Once initiated, additional mechanisms limit the duration of this amplifying cascade and restore the various molecules to their inactivated states. The protein arrestin, for instance, blocks the ability of activated rhodopsin to activate transducin, and facilitates the breakdown of activated rhodopsin. The all-trans retinal then dissociates from the opsin, diffuses into the cytosol of the outer segment, is converted to all-trans retinol and is transported out of the outer segment and into the pigment epithelium, where appropriate enzymes ultimately convert it to 11-cis retinal. After it is transported back into the outer segment, the 11-cis retinal recombines with opsin in the receptor disks. The recycling of rhodopsin is critically important for maintaining the light sensitivity of photoreceptors. Even under intense levels of illumination, the rate of regeneration is sufficient to maintain a significant number of active photopigment molecules. Functional Specialization of the Rod and Cone Systems the two types of photoreceptors, rods and cones, are distinguished by shape (from which they derive their names), the type of photopigment they con- Vision: the Eye 241 (A) Rod Disks (B) Cone Figure 10. Although generally similar in structure, rods (A) and cones (B) differ in their size and shape, as well as in the arrangement of the membranous disks in their outer segments. Outer segment Cytoplasmic space Plasma membrane Cilium Mitochondria Outer segment Inner segment Nucleus Inner segment Synaptic terminal Synaptic vesicles Synaptic terminal tain, distribution across the retina, and pattern of synaptic connections (Figure 10. These properties reflect the fact that the rod and cone systems (the receptors and their connections within the retina) are specialized for different aspects of vision. The rod system has very low spatial resolution but is extremely sensitive to light; it is therefore specialized for sensitivity at the expense of resolution. Conversely, the cone system has very high spatial resolution but is relatively insensitive to light; it is therefore specialized for acuity at the expense of sensitivity. The properties of the cone system also allow humans and many other animals to see color. The range of illumination over which the rods and cones operate is shown in Figure 10. The difficulty of making fine visual discriminations under very low light conditions where only the rod system is active is a common experience. The problem is primarily the poor resolution of the rod system (and, to a lesser degree, the fact that there is no perception of color in dim light because the cones are not involved to a significant degree). Although cones begin to contribute to visual perception at about the level of starlight, spatial discrimination at this light level is still very poor. As illumination increases, cones become more and more dominant in determining what is seen, and they are the major determinant of perception under relatively bright conditions such as normal indoor lighting or sunlight. The contributions of rods to vision drops out nearly entirely in socalled photopic vision because their response to light saturates-that is, the membrane potential of individual rods no longer varies as a function of illumination because all of the membrane channels are closed (see Figure 10. Mesopic vision occurs in levels of light at which both rods and cones contribute-at twilight, for example. From these considerations it should be clear that most of what we think of as normal "seeing" is mediated by the cone system, and that loss of cone function is devastating, as occurs in 242 Chapter Ten -6 -4 Starlight -2 Luminance (log cd/m-2) 2 0 Indoor lighting 4 Sunlight 6 8 Moonlight Luminance of white paper in: No color vision Poor acuity Visual function Scotopic Absolute threshold Cone threshold Mesopic Good color vision Best acuity Photopic Rod saturation begins Best acuity 50% bleach Indirect ophthalmoscope Damage possible Figure 10. Cones begin to contribute to perception at about the level of starlight and are the only receptors that function under relatively bright conditions. People who have lost cone function are legally blind, whereas those who have lost rod function only experience difficulty seeing at low levels of illumination (night blindness; see Box B). Differences in the transduction mechanisms utilized by the two receptor types is a major factor in the ability of rods and cones to respond to different ranges of light intensity. For example, rods produce a reliable response to a single photon of light, whereas more than 100 photons are required to produce a comparable response in a cone. Rather, the change in current produced by single photon capture in cones is comparatively small and difficult to distinguish from noise. Another difference is that the response of an individual cone does not saturate at high levels of steady illumination, as does the rod response.
Purchase on line female viagra. Las Vegas women's healthcare - University of Nevada School of Medicine.